
文章信息
● 原名:Global health risks lurking in livestock resistome
● 译名:畜禽抗性组潜藏的全球健康风险
● 期刊:Science Advances
● IF:12.5
● 通讯作者:谷洁、James Tiedje、钱勋
● 通讯单位:西北农林科技大学资源环境学院
● 论文DOI:10.1126/sciadv.adt8073
导读
畜牧业每年消耗全球超过70%的抗生素,使其粪便成为人为影响下抗生素抗性基因(ARGs)的重要载体。目前,畜禽抗性组的全球分布模式、驱动机制及向临床传播的潜力尚未得到充分评估。在该研究中,研究者分析了来自26个国家的4017份畜禽粪便宏基因组,构建了畜禽ARGs和宏基因组组装基因组的综合目录。畜禽抗性组是已知(2291种亚型)和潜在ARGs(3166种亚型)的重要储存库,且与人类抗性组高度关联。研究者描绘畜禽抗性组的全球分布模式及临床关键ARGs的流行情况,强调农场管理和人类抗生素管理在塑造畜牧业抗性组中的作用。研究者通过整合迁移潜力、临床意义和宿主致病相关性构建风险评估框架,对高风险畜牧业ARGs进行优先排序,生成可预测的畜禽抗性组风险全球地图,有望为研究和政策制定提供指导。
实验设计
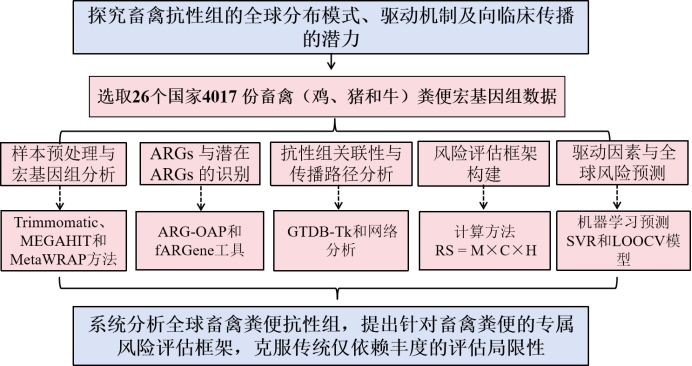
主要内容
1. 庞大的畜禽粪便泛抗性组及其与人类和环境抗性组的关联性
研究者从4017份粪便宏基因组中鉴定出2291种ARG亚型,这些亚型可能赋予对30类抗生素的抗性(图1A)。所有三种粪便类型均携带开放泛抗性组,其中鸡粪的ARG多样性最高(2107种亚型),其次是猪粪(1779种亚型)和牛粪(1546种亚型)(图1B)。平均而言,每份鸡粪样本携带246种ARG亚型和每个细胞4.33个拷贝,高于猪粪(203种亚型和每个细胞3.12个拷贝)和牛粪(129种亚型和每个细胞0.89个拷贝)(图1C、D)。通常,ARG多样性高与抗性组丰度高相关,尽管部分抗性组丰度高的样本显示相对较低的ARG多样性(图S1A)。高度流行的ARG也表现出高丰度。值得注意的是,38种ARG在超过90%的畜禽粪便样本中被检测到(图1E和表S4),分别占鸡粪、猪粪和牛粪抗性组丰度的47%、63%和80%(图S1B)。相反,大多数ARG(2291种中的1887种)仅在不足10%的样本中被检测到(图1E)。抗性组特征显著受宿主动物影响[置换多元方差分析(PERMANOVA),P < 0.001;图S1C]。
宿主相关基质(畜禽粪便、人类粪便和污水)中的抗性组特征与自然环境(土壤、沉积物和水)显著不同(PERMANOVA,P < 0.001;图1F)。畜禽粪便与人类粪便的抗性组特征重叠程度高于污水与人类粪便的重叠程度。除污水外,畜禽粪便的ARG多样性最高(图1C),其中鸡粪和猪粪抗性组的ARG丰度在所有ARG储存库中最高(图1D)。鸡粪和猪粪宏基因组的平均抗性组丰度是人类粪便的2.0倍、污水的2.5倍和土壤的18.3倍。人类粪便中发现的几乎所有ARG也存在于畜禽粪便中,人类粪便分别与鸡粪、猪粪和牛粪共享1250、1114和976种ARGs(图1G)。
ARGs在不同环境中的传播可通过共栖于这些环境的同一细菌物种实现。因此,研究者通过计算两个栖息地共享的同一属ARGs评估二者抗性组的连通性。畜禽粪便与人类粪便和污水的抗性组连通性最强,分别可能传播1761种和1626种ARG亚型(图1F和图S2A)。畜禽粪便中的大肠杆菌(Escherichia)、克雷伯氏菌(Klebsiella)和沙门氏菌(Salmonella)是最重要的载体,可分别介导240种、123种和110种畜禽ARGs向其他环境的传播(图S2B)。
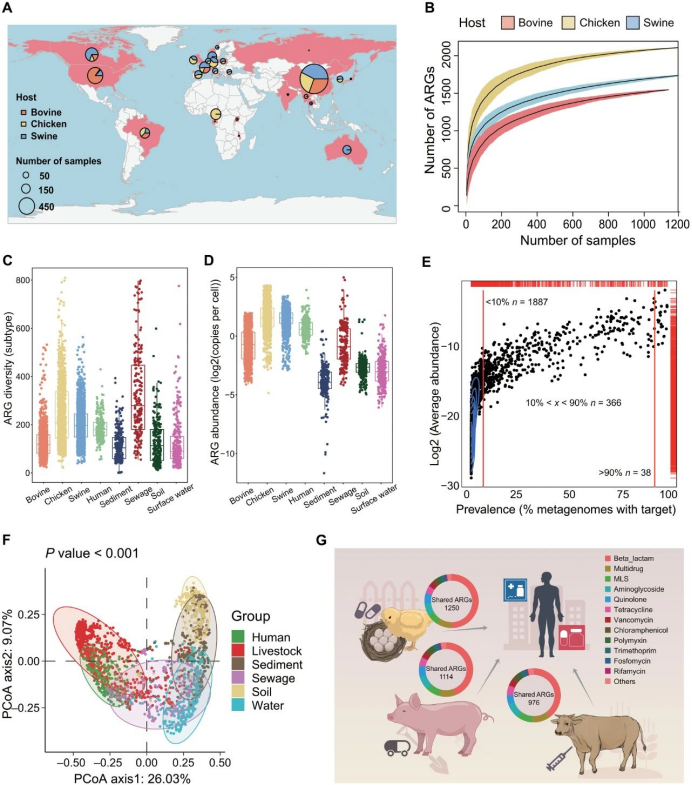
图1. 畜禽粪便、人类粪便及典型ARG储存库中的ARGs。(A) 4017份畜禽粪便宏基因组的地理分布。(B) 畜禽粪便泛抗性组。(C) ARG多样性及(D) ARG丰度在畜禽粪便与典型ARG储存库中的对比(每个点代表一个宏基因组)。(E) 畜禽粪便样本中ARG的平均丰度与普遍性(顶部和右侧红线表示ARG密度分布)。(F) 主坐标分析(PCoA)展示不同环境抗性组特征的差异(每种动物粪便随机选取200个宏基因组绘制抗性组特征)。(G) 人类粪便与畜禽粪便共享的ARGs。
2. 畜禽粪便是潜在抗生素抗性基因的重要储存库
潜在ARGs指现有方法和数据库无法检测到但可能赋予抗生素抗性的基因。相比之下,通过标准ARGs-OAP方法检测到的ARGs称为已确定ARGs。研究者在畜禽粪便中预测到共3166种潜在ARGs(图2A),数量超过已确定ARGs的多样性(2291种亚型),其中氨基糖苷类和β-内酰胺类抗性基因占主导。除牛粪外,鸡粪和猪粪中潜在ARGs的丰度显著低于已确定ARGs(P < 0.001;图2B)。大多数潜在ARGs仅在特定类型的粪便中检测到。选取3种在畜禽粪便中丰度高、流行广且具有广宿主范围的潜在ARGs进行抗性表型验证。当在大肠杆菌(Escherichia coli)TOP10中表达时,AAC(6′)-c1和APH(6)-c2可赋予对庆大霉素(32 μg/ml)的抗性,erm-c1可赋予对红霉素(64 μg/ml)的抗性(图S3)。
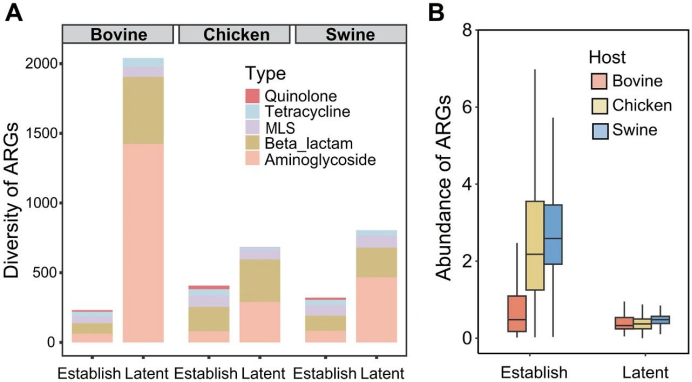
图2. 畜禽粪便中的潜在与已确定ARGs。(A) 潜在与已确定ARGs的多样性比较。(B) 已确定与潜在ARGs的丰度比较。潜在ARGs通过fARGene预测,已确定ARGs通过ARGs-OAP鉴定。
3. 畜禽ARGs的广宿主范围与致病性关联
研究者从畜禽粪便宏基因组中共构建123872个宏基因组组装基因组(MAGs);其中12069个被鉴定为携带563种ARG的宿主(图3)。这些ARG宿主属于28个细菌门、55个纲、129个目、276个科、1010个属和1818个种。假单胞菌门(Pseudomonadota)、芽孢菌门(Bacillota)和拟杆菌门(Bacteroidetes)是畜禽ARG的主要宿主(图3)。不同门宿主携带的ARG类型存在差异,Pseudomonadota主要携带多重耐药基因,占总ARG多样性的71%;Bacillota携带32%的大环内酯-林可酰胺-链阳霉素(MLS)类、21%的四环素类和15%的氨基糖苷类抗性基因;Bacteroidetes携带32%的MLS类和29%的β-内酰胺类抗性基因。
大约34%携带ARG的MAGs包含编码对至少三类抗生素抗性的基因(图3)。与其他ARG宿主相比,肠杆菌科(Enterobacteriaceae)和葡萄球菌科(Staphylococcaceae)携带更多ARGs。这些富含ARG的Enterobacteriaceae和Staphylococcaceae主要存在于亚洲的猪粪样本中。此外,放线菌门(Actinomycetota)的一个分支携带多种多样的ARGs,其中大部分在欧洲的鸡粪样本中被鉴定(图3)。研究者在人类病原体中共检测到340种ARGs,其中多重耐药(28%)、β-内酰胺类(20%)和MLS(11%)抗性基因最为常见。潜在病原体携带的ARGs数量(每个MAG 17.8个拷贝)显著多于非致病菌(每个MAG 1.6个拷贝)(P < 0.001;图S4)。
研究者共鉴定出174个抗生素抗性基因(ARGs),这些基因至少存在于三个细菌门类、249个科和930个属中(图S5)。MLS类抗性基因Lnu(C)由214个细菌属携带,四环素类抗性基因tet(44)存在于111个属中(图S6)。在广宿主范围的ARGs(存在于≥3个门中)中,174种里有105种与移动遗传元件(MGEs,如tnp、rep和intI)相关联。
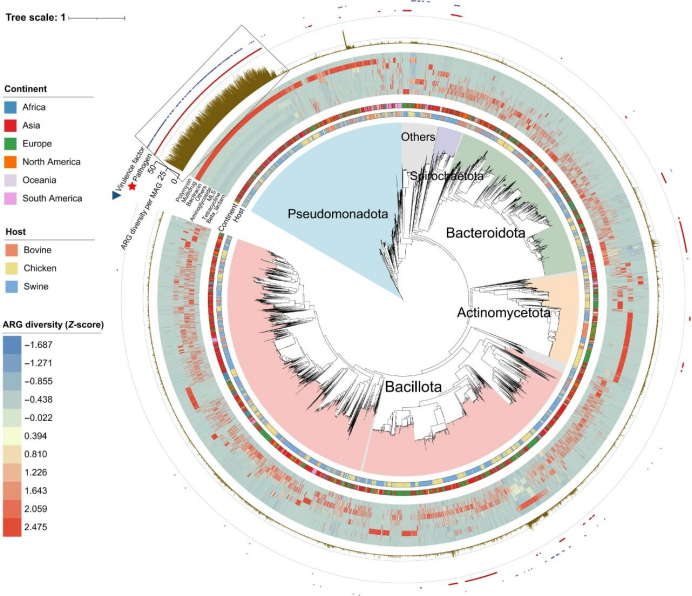
图3. 基于畜禽宏基因组构建的MAGs系统发育树。系统发育树的每个分支代表一个由宏基因组构建的MAG。背景颜色表示不同的细菌门。从内向外数的第一圈和第二圈分别表示每个MAG所属的大陆和动物宿主。热图展示每个MAG携带的不同ARG类别的比例。柱状图显示每个MAG携带的ARG多样性。从外向内数第二圈的蓝色三角形表示该MAG携带毒力因子。框内显示,携带高多样性ARG的MAG主要归类于Enterobacteriaceae。
4. 畜禽粪便中ARGs风险优先级评估框架
为构建一个稳健、功能完善且准确的风险评估框架,研究者整合三个维度的六个指标:迁移性(M)(表S5)、临床重要性(C)(表S6)和宿主致病相关性(H)(表S9)。应用该框架时,可为宏基因组中的单个ARG分配单一风险评分(RS)(图4C)。该框架被用于对从畜禽粪便MAGs中检测到的563种ARG,以及从畜禽粪便分离株基因组中鉴定出的173种ARG进行优先级排序(图4A和表S10)。这736种ARG代表最常观测到的ARG(图S7A),且在样本中共同占抗性组丰度的95%以上(图S7B)。在分类的736种ARG中,约52.6%与MGEs相关。多重耐药基因和万古霉素耐药基因主要位于染色体上,而sul1、TEM-1、OXA-1和QnrS1则主要位于质粒上。ARG宿主与致命病原体之间的进化距离计算为H1值,其与表示ARG宿主毒力能力的H2值呈现相似趋势(图4B)。大多数MLS和万古霉素耐药基因的H1值较低,而其他抗生素类别的ARG的H1值差异显著。按H1值排序的前三分之一的ARG主要包括β-内酰胺类(87种)、多重耐药类(52种)、氨基糖苷类(33种)、多粘菌素类(16种)和喹诺酮类(13种)耐药基因。氯霉素耐药基因catII和β-内酰胺类耐药基因DHA-16显示出最高的H1值,表明其宿主细菌具有高度致病相关性。
研究者将迁移性(M)、临床重要性(C)和宿主致病相关性(H)的乘积定义为风险评分(RS),并将畜禽ARGs划分为四个风险等级(图4C)。R1级(RS≥1)包含44种ARGs,主要由β-内酰胺类耐药基因(34种)组成,辅以氨基糖苷类(3种)、MLS类(1种)、喹诺酮类(4种)、链丝菌素类(1种)和磺胺类(1种)耐药基因(图4D)。R2级(1>RS≥0.2)、R3级(0.2>RS>0)和R4级(RS=0)分别包含159、265和268种ARGs。值得注意的是,RS=0并不表示无风险,而是因迁移性或临床相关性证据不足导致的相对低风险。β-内酰胺类耐药基因在除R4级外的所有风险等级中分布均匀;多粘菌素类耐药基因主要分布于R2级;万古霉素类耐药基因则主要落在R3级(图S7C)。研究者将样本的抗性组风险评分(RS)定义为每个ARG的RS与其丰度的累积乘积。通过量化三种畜禽粪便类型的抗性组风险评分,研究者发现鸡粪(0.94±1.41)风险最高,其次是猪粪(0.39±0.33),牛粪风险最低(0.09±0.11)(图S7D)。
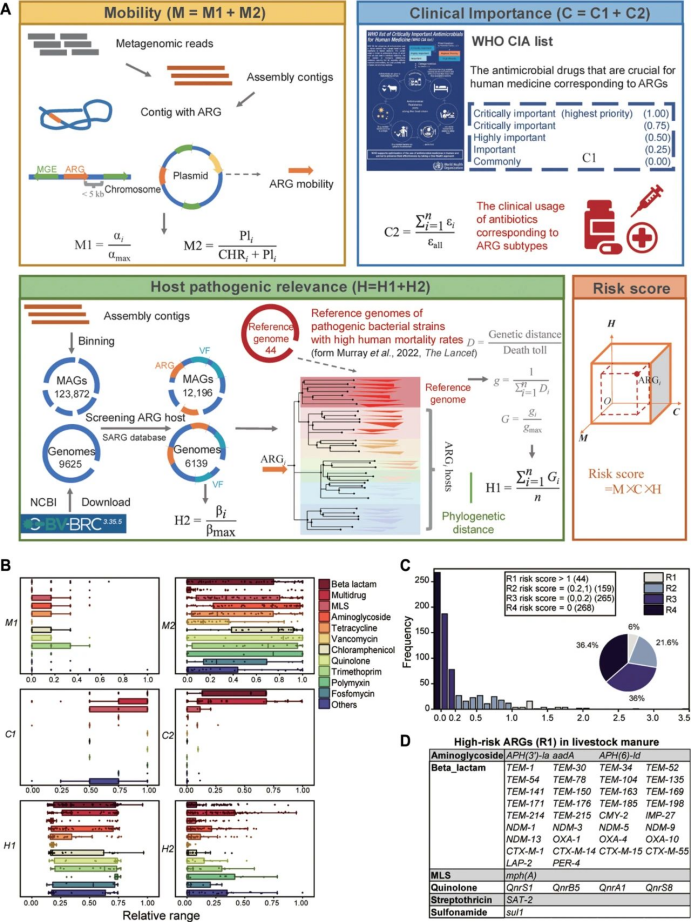
图4. 畜禽粪便中ARGs风险评估框架。(A)展示风险评估框架构建概念与流程的示意图。该框架基于三个维度的测量:迁移性(M)(表S5)、临床重要性(C)(表S6)和宿主致病相关性(H)(表S9)。(B)不同类型ARGs风险指标的相对范围。相对范围表示各指标的计算值,具体数据详见表S5-S10。(C)按四个风险等级(R1、R2、R3、R4)分类的ARGs分布。饼图展示所有ARGs的分布,柱状图显示风险评分分布。阈值0表示ARGs在至少一个风险维度得分为零的情况,阈值1和0.2基于柱状图所示风险评分分布经验性确定。(D)畜禽粪便中高风险ARGs(R1)清单。
5. 全球畜禽耐药组的显著差异模式
研究者观察到不同大陆和国家间畜禽耐药组的丰度、多样性和特征存在差异(图S8)。亚洲猪粪耐药组的多样性(246种ARGs)和丰度(每个细胞3.93个拷贝)显著高于其他大陆(P < 0.01;图S9),尤其是氨基糖苷类、MLS类和四环素类抗性基因的丰度(图S10)。欧洲样本中鸡粪耐药组的多样性最高(314种ARGs)。值得注意的是,非洲鸡粪中β-内酰胺类抗性基因的丰度和北美鸡粪中MLS抗性基因的丰度均高于其他大陆(图S10)。北美牛粪的耐药组丰度(每个细胞1.3个拷贝)和ARG多样性(160种ARGs)最高,其丰度值分别为亚洲和欧洲的3.3倍和5.1倍。北美牛粪耐药组的高丰度主要由高丰度的四环素类(如tetW和tetQ)和MLS抗性基因(如lnuC和ermB)驱动。
鸡粪耐药组丰度的最高值和最低值均出现在欧洲国家,其中最高值见于波兰(每个细胞5.26个拷贝)、保加利亚(4.95个拷贝)和西班牙(4.57个拷贝),最低值见于德国(0.61个拷贝)(图5A)。猪粪耐药组的最高丰度和多样性出现在中国(每个细胞3.98个拷贝,250种ARGs),显著高于所有其他国家(P < 0.01,方差分析;图5A、B)。在欧洲国家中,意大利、西班牙和保加利亚的猪粪耐药组丰度高于其他欧洲国家。挪威的猪粪耐药组丰度(每个细胞0.50个拷贝)和ARG多样性(101种亚型)最低。在提供牛粪宏基因组的六个国家中,加拿大的牛粪耐药组丰度最高(每个细胞2.50个拷贝),其次是美国(1.07个拷贝)。中国的牛粪耐药组丰度为加拿大的15.2%。
研究者调查六种临床关键ARGs及其变体(CTX-M、KPC、NDM、VIM、mcr和tetX)的宿主和全球流行模式,以及44种(R1)高风险ARGs的全球流行模式(图5C)。这六种ARGs的总体流行率低于10%。β-内酰胺类抗性基因KPC、VIM和NDM在中国的流行率最高,而tetX在多个国家广泛分布,其中tetX2(79.5%)和tetX6(66.8%)表现出最高的流行率。多粘菌素mcr抗性基因在欧洲国家更常见,具体而言,保加利亚和丹麦的mcr流行率高于其他国家。CTX-M在中国的流行率最高,其次是德国、意大利和缅甸。
研究者分析六个主要畜禽生产国抗性组丰度的时间变化(图5D)。在美国,牛粪抗性组丰度自2013年起持续下降,并于2015-2018年趋于稳定。相比之下,中国牛粪抗性组丰度呈上升趋势,并于2018年达到峰值。除西班牙(2021年)和德国(2022年)外,未观察到鸡粪抗性组丰度随时间的下降趋势。2013-2019年,中国猪粪抗性组丰度总体呈下降趋势。2016-2018年,美国猪粪抗性组丰度显著降低。德国和法国猪粪的抗性组丰度多年来保持稳定。
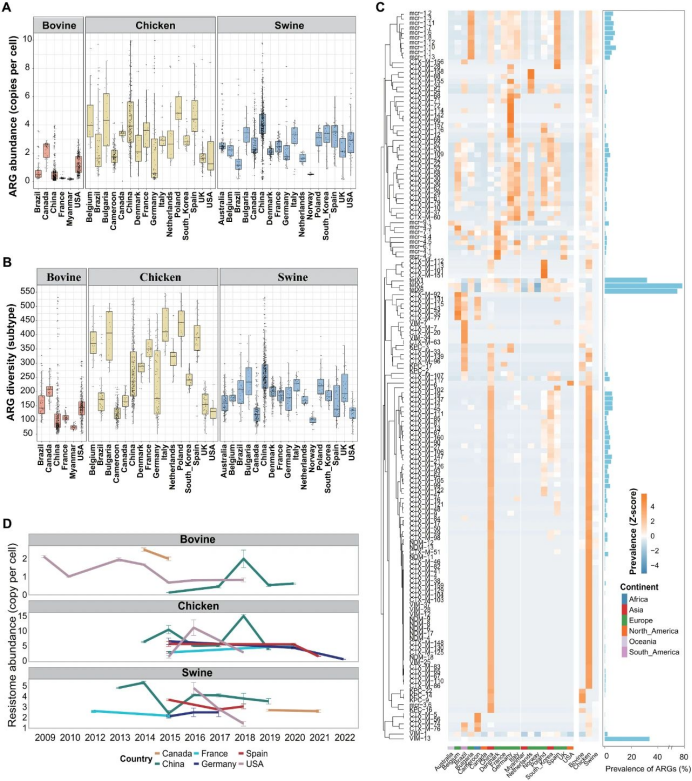
图5. 全球畜禽耐药组分布模式。(A) 三种动物粪便耐药组丰度及(B) ARG多样性在不同国家的差异。(C) 临床关键抗生素抗性基因在各大洲及国家的流行情况,右侧柱状图展示畜禽粪便中ARG的总体流行率。(D) 六大主要畜禽生产国耐药组丰度的时间变化趋势。
6. 全球畜禽粪便耐药组风险分布图
研究者采用支持向量回归(SVR)与留一法交叉验证(LOOCV)的协同方法,评估潜在驱动因素对畜禽耐药组RS的贡献(图6A)。基于均方误差(MSE;图6B),保留16个变量并构建预测模型。该模型在预测畜禽耐药组RS方面表现良好(图6C)。畜禽密度、畜禽及人类抗菌药物使用量、畜禽生产量是畜禽耐药组RS的主要决定因素(图6D)。畜禽粪便中耐药组的丰度与人均国内生产总值(GDP)呈负相关,但与农场抗菌药物使用量呈正相关(图S11)。
利用该模型,研究者生成2022年鸡粪和猪粪RSs的预测世界地图(图6E)。与耐药组特征一致,鸡粪和猪粪的耐药组风险呈现出明确的大陆和国家模式。非洲的鸡粪RSs预测值最高,而欧洲的RS预测值最低。美洲猪粪的ARG风险低于非洲和欧洲。该地图概述畜禽粪便中的抗生素耐药威胁,对监测工作至关重要。
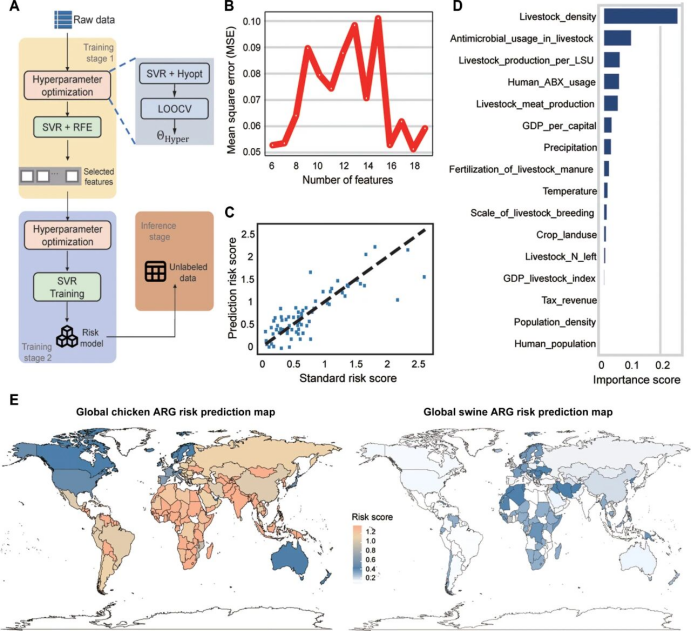
图6. 畜禽粪便耐药组风险评分预测全球地图。(A) 用于预测全球畜禽耐药组风险评分的机器学习示意图。(B) 均方误差(MSE)评估。(C) 预测风险评分与真实风险评分的相关性。(D) 用于预测畜禽耐药组风险评分的前16个因素。各因素全称见补充数据。(E) 2022年全球鸡粪和猪粪耐药组风险评分预测地图。色标表示绝对风险评分。白色填充国家因无畜禽生产未纳入风险预测。
讨论
该研究揭示畜禽粪便是一个开放的泛抗性组储存库,其ARG多样性高于此前认知,涵盖几乎所有已知ARG及大量未表征的潜在ARG。畜禽抗性组与人类抗性组高度关联,且其每个细胞携带的ARG数量超过人类粪便、污水和土壤等典型ARG热点环境。Enterobacteriaceae和Staphylococcaceae等肠道共生菌可介导超过240种ARG在畜禽与人类间的传播。畜禽抗性组中相当一部分ARG具有广泛宿主范围,部分ARG宿主可在多样生态位中存活。因此,畜禽粪便的不当处置可能会促进ARG在广泛生态系统中的传播与定殖。鉴于畜禽抗性组的高致病相关性及其与人类、自然环境的相互关联,研究者建议将畜禽粪便作为废水的替代“哨兵环境”,为ARG增殖及公共卫生系统风险恶化提供早期预警。
畜禽粪便中的抗性组特征受宿主动物影响,其多样性和丰度呈现层级差异:鸡>猪>>牛。这与先前研究一致,可能源于不同动物类型肠道微生物组和抗生素使用的差异。例如,鸡和猪的人口校正单位(PCU)抗生素消耗量远高于牛。仅38种ARG存在于所有类型的畜禽粪便中。它们代表常见肠道细菌携带的组成型ARG,贡献抗性组丰度的主要部分。相反,大多数畜禽ARG在样本中表现出低流行率和高变异性。这些ARG被定义为“可变ARG”,代表从环境中富集和获得的ARG。可变ARG的高多样性强调养殖方式和抗生素暴露对畜禽抗性组的重大影响,并表明通过识别和实施合理的农场管理,有望缓解畜禽养殖中的抗生素耐药性。
识别高风险ARGs对畜禽养殖环境中抗生素耐药性的监测至关重要。先前评估ARG风险的方法未考虑不同生态系统中ARGs在丰度、流行率、宿主范围和迁移性上的差异。该研究通过整合畜禽抗性组的独特特征,开发针对畜禽抗性组的风险评估框架。该概念与方法亦可应用于其他环境中ARGs的风险优先级评估。该研究的框架整合迁移潜力、临床重要性和宿主致病相关性,将畜禽粪便中的736种ARG划分为四个风险等级。值得注意的是,研究者提出利用ARG宿主与导致全球主要死亡的病原体之间的进化距离,为评估不同ARG的临床影响提供实用方法。研究者识别出44种高风险ARG(R1级),这些ARG更易转移至致命病原体并可能削弱抗生素治疗效果。部分识别的高风险ARG(如TEM、NDM、OXA和CTX-M),已被报道可从畜禽环境传播至临床环境并引发严重临床后果。此外,基于每个ARG的风险评分,研究者提出一种评估样本整体抗性组风险的定量方法。研究发现,鸡粪的抗性组风险高于猪粪,显著高于牛粪,这可能与家禽养殖中更高的抗生素使用量、更快的微生物周转及集约化养殖方式相关。研究者承认该框架存在一定局限性。例如,其未包含未在研究者宏基因组数据集中组装的部分ARGs,且从组装质量差的ARGs推断的宿主和迁移信息需进一步验证。值得注意的是,基于宏基因组数据的风险评估框架在确定表型特征(如致病性,其依赖于菌株和环境)和耐药表型,以及质粒的转移性和宿主范围方面存在局限。对ARG的精确风险评估需要菌株水平的临床验证和实验数据,这超出当前评估框架的范围。
据研究者所知,该研究是目前对畜禽耐药组最全面的调查,覆盖14年数据和26个不同收入水平国家及9个主要畜禽生产国。畜禽耐药组特征呈现显著的大陆和国家差异。作为全球最大的生猪生产国(占全球生猪存栏量50%以上),中国在猪粪中的细菌丰度、多样性和风险评分均高于所有其他国家。同样,作为主要牛肉生产国,美国和巴西的牛粪耐药组丰度与多样性(除加拿大外)显著高于其他国家。这些发现强调需重点关注主要畜禽生产国的耐药组风险。尽管欧盟在畜禽养殖中限制抗生素使用的历史悠久,但各国鸡粪耐药组的多样性和丰度仍存在差异。特别是保加利亚和西班牙的鸡粪、猪粪耐药组丰度与风险显著高于其他欧洲国家(如挪威、丹麦和德国),这与这些国家的畜禽抗生素使用量一致。亚洲畜禽粪便中氨基糖苷类(如APH(3′)-IIIa和aad(6))和MLS类(如lnuC和erm)抗性基因的丰度远高于其他国家,而北美牛粪中需更关注MLS(如lnuC和ermB)和四环素类(如tetW和tetQ)抗性基因。非洲畜禽耐药组在本研究中覆盖较少,但研究者观察到喀麦隆鸡粪中β-内酰胺类(如CfxA和TEM)抗性基因丰度较高,凸显非洲需加强ARG监测。值得欣慰的是,限制农场抗生素使用对缓解抗生素耐药性产生积极效果。中国猪粪耐药组丰度自2014年起持续下降,可能得益于其限制抗生素促生长使用的法规。类似地,美国猪粪耐药组的丰度与多样性在2016-2018年间快速降低,可能与2017年《兽医饲料指令》政策的实施相关。
通过机器学习方法,研究者识别出畜禽及人类抗生素使用量、畜禽生产规模和密度在驱动全球畜禽耐药组中的重要作用。畜禽养殖中抗生素使用的特征仍是耐药组丰度和风险的主要决定因素,但无法完全解释观察到的变异性。与先前研究一致,该研究的结果证实耐药组丰度与人居GDP的负相关关系,以及其与畜禽生产密度和规模的正相关关系。最终,该研究提供全球预测地图以洞察全球畜禽耐药组风险热点。此外,该研究生成畜禽粪便ARGs的全面目录,包括ARG序列、携带ARG的contig和MAGs。该目录不仅概述本时期畜禽粪便中的耐药组特征,还为研究耐药组传播、健康风险及评估实践提供宝贵资源。然而,研究者承认这些资源依赖短读长宏基因组数据,存在潜在遗漏低丰度ARGs及无法提供ARG绝对丰度测量等局限性。
资讯来源:微信公众号”生命科学进展“推文《Sci Adv│西北农林谷洁/钱勋教授团队揭示养殖环境耐药基因的传播机制与全球健康风险》
阅读推文
阅读原文